Spore structure
The study of zonate-camerate spores through Transmission Electron Microscopy (TEM) (e.g. Archangelsky and Césari, 1990; Raine et al. 1988) has led to a better understanding of their internal structure and affinities. However TEM is expensive, time consuming, involves concentration on a small number of specimens, and is often inconsistent with practical palynological study which is still overwhelmingly carried out by light microscopy. In this article I show that light microscopy of well-preserved, and particularly of broken specimens, can reveal much about the internal structure of specimens of Vallatisporites.

Vallatisporites-type spores
Vallatisporites-type spores are common in the Carboniferous-Permian sedimentary rocks of the Arabian peninsula and are often associated with fluvial or lacustrine glacial facies of the 2165 and OSPZ2 biozones (Penney et al. 2008, Stephenson et al. 2003, Stephenson 2004). The spores are large, often reaching more than 100 microns in diameter. They are trilete, rounded triangular, to almost circular. Their internal and external structure is complex, with grana and spinae on the proximal sides and sometimes complex structures on the distal side that appear to support the zona, perhaps to offer greater strength to the zona, which itself likely had a role in dispersal and perhaps in positioning during germination.
Along with other specimens of the genus Vallatisporites, these spores usually have a row of ‘pits’ or ‘intra-zona’ vacuoles, with uncertain function, close to the inner margin of the zona. Within the enclosing structure of the spore and zona is an inner body that appears relatively simple and unornamented. This body often does not appear to completely fill the internal cavity of the spore so that a 5 µm or so wide space between the margin of the inner body and the inner margin of spore cavity can be seen when viewed in proximodistal orientation. The concentric position of the inner body within the spore cavity tends to suggest adpression of the exinal layers at the proximal and/or distal poles.

Some of these characters are shown in Fig. 1 which is a very well preserved specimen. However although much can be gained from studying ‘perfect’ specimens, a lot can also be gained by light microscopy analysis of broken specimens that nevertheless otherwise show aspects of good preservation.
In Fig. 2 a broken specimen that is otherwise quite well preserved shows some of the structural elements better than a complete specimen. This specimen seems to indicate aspects of the inner body and of the margins of the proximal part of the outer spore structure close to the zona. The inner body is clearly punctate rather than completely smooth. It is also quite a bit smaller than the encasing spore body. It appears not to have laesurae (a trilete mark). The proximal part of the outer spore structure close to the zona also seems to be punctate rather than smooth. This layer of exine, besides being punctate, also seems relatively thin and delicate, at least in comparison with the rather rigid zona.

In Fig. 3 another broken specimen, showing an intexinal body that is clearly quite thick walled and punctate is close to being liberated from the spore cavity. The distal exine is seen to be coarsely ornamented but relatively thin in between ornament elements.
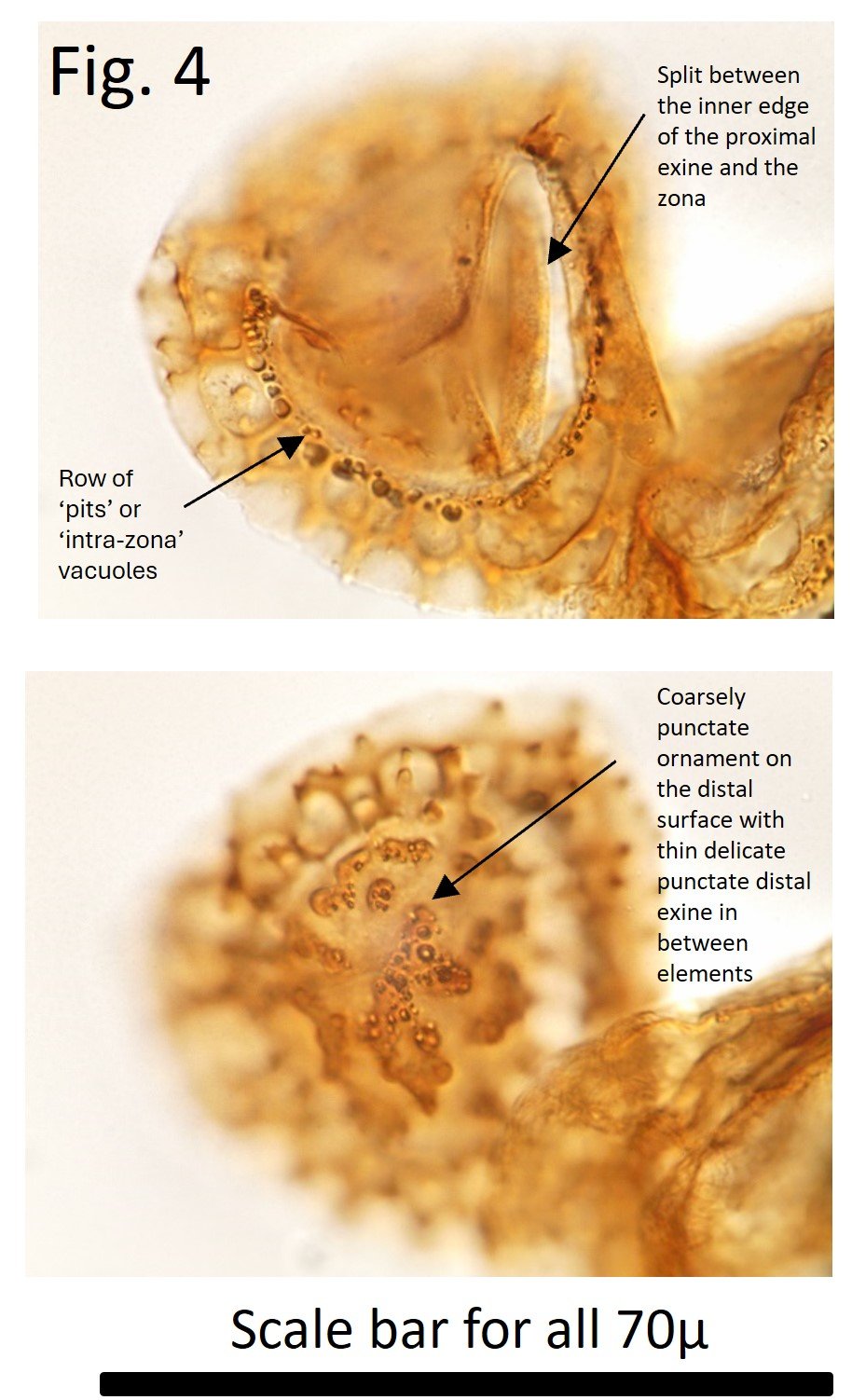
Fig. 4 shows an only slightly damaged specimen with a well displayed row of ‘pits’ or ‘intra-zona’ vacuoles and a split between the inner edge of the proximal exine and the zona. On the distal surface is very coarsely punctate ornament with thin delicate punctate distal exine in between ornamemt elements. Again the delicacy of this exine, despite the thick ornamentation, will tend to make this part of the spore relatively weak. Fig. 5 again shows a punctate inner body, thin delicate punctate exine that extends beyond inner body on the proximal side, and some coarsely punctate ornament on the distal surface. Perhaps the complex distal structures under the zona seen in this figure, and Fig. 1 provide support to the zona - and structural rigidity - ensuring that spores remained intact for reproductive purposes.

Finally Fig. 6 shows the parts of Vallatisporites-type spores with only the proximal exine and zona, and distal exine and zona preserved respectively, so each specimen lacks an intexinal body, and one layer of external exine - proximal or distal. Although not common, these broken specimens allow very detailed study of these different surfaces without the encumbrance of other layers that may obscure detail.

References
Archangelsky, S. and Césari, S. 1990. Ultrastructura de la exina en ejemplares Carboniferos de Lundbladispora (Licofita), La Rioja, Argentina. Ameghiniana, v. 27, p. 131-139.
Raine, J. I., de Jersey, N. J. and Ryan, K. G. 1988. Ultrastructure and lycopsid affinity of Densoisporites psilatus (de Jersey) comb. nov. from the Triassic of New Zealand and Queensland. Mem. Ass. Australas. Palaeontols. v. 5, p. 79-88.
Penney, R. A., Al Barram, I., and Stephenson, M. H., 2008. A high resolution palynozonation for the Al Khlata Formation (Pennsylvanian to Lower Permian), South Oman. Palynology, v. 32, p. 213-231.
Stephenson, M.H. Early Permian 2004. Spores from Saudi Arabia and Oman. In Al-Husseini, M (ed.) Carboniferous, Permian and Early Triassic Arabian stratigraphy, GeoArabia Special Publication 3, Gulf PetroLink, Manama, Bahrain, p. 185-215.
Stephenson, M. H., Osterloff, P. L. and Filatoff, J. 2003. Integrated palynological biozonation of the Permian of Saudi Arabia and Oman: progress and problems. GeoArabia, v. 8, p. 467-496.